
Фотосинтез - это синтез органических соединений в листьях зеленых растений из воды и углекислого газа атмосферы с использованием солнечной (световой) энергии, адсорбируемой хлорофиллом в хлоропластах.
Благодаря фотосинтезу происходит улавливание энергии видимого света и превращение ее в химическую энергию, сохраняемую (запасаемую) в органических веществах, образуемых при фотосинтезе.
Датой открытия процесса фотосинтеза можно считать 1771 г. Английский ученый Дж. Пристли обратил внимание на изменение состава воздуха вследствие жизнедеятельности животных. В присутствии зеленых растений воздух вновь становился пригодным как для дыхания, так и для горения. В дальнейшем работами ряда ученых (Я. Ингенгауз, Ж. Сенебье, Т. Соссюр, Ж.Б. Буссенго) было установлено, что зеленые растения из воздуха поглощают С0 2 , из которого при участии воды на свету образуется органическое вещество. Именно этот процесс в 1877 г. немецкий ученый В. Пфеффер назвал фотосинтезом. Большое значение для раскрытия сущности фотосинтеза имел закон сохранения энергии, сформулированный Р. Майером. В 1845 г. Р. Майер выдвинул предположение, что энергия, используемая растениями, - это энергия Солнца, которую растения в процессе фотосинтеза превращают в химическую энергию. Это положение было развито и экспериментально подтверждено в исследованиях замечательного русского ученого К.А. Тимирязева.
Основная роль фотосинтезирующих организмов:
1) трансформация энергии солнечного света в энергию химических связей органических соединений;
2) насыщение атмосферы кислородом;
В результате фотосинтеза на Земле образуется 150 млрд. т. органического веществаи выделяется около 200 млрд. т свободногокислородав год. Он препятствует увеличению концентрацииCO2в атмосфере, предотвращая перегрев Земли (парниковый эффект).
Созданная фотосинтезом атмосфера защищает живое от губительного коротковолнового УФ-излучения (кислородно-озоновый экран атмосферы).
В урожай сельскохозяйственных растений переходит лишь 1-2% солнечной энергии, потери обусловлены неполным поглощением света. Поэтому имеется огромная перспектива повышения урожайности благодаря селекции сортов с высокой эффективностью фотосинтеза, созданию благоприятной для светопоглощения структуры посевов. В связи с этим особенно актуальными становятся разработка теоретических основ управления фотосинтезом
Значение фотосинтеза гигантское. Отметим лишь, что он поставляет топливо (энергию) и атмосферный кислород, необходимые для существования всего живого. Следовательно, роль фотосинтеза является планетарной.
Планетарность фотосинтеза определяется также тем, что благодаря круговороту кислорода и углерода (в основном) поддерживается современный состав атмосферы, что в свою очередь определяет дальнейшее поддержание жизни на Земле. Можно сказать далее, что энергия, которая запасается в продуктах фотосинтеза, есть по существу основной источник энергии, которым сейчас располагает человечество.
Суммарная реакция фотосинтеза
СО 2 +Н 2 О = (СН 2 О) + О 2 .
Химию фотосинтеза описывают следующими уравнениями:

Фотосинтез – 2 группы реакций:
световая стадия (зависят от освещенности)
темновая стадия (зависит от температуры).
Обе группы реакций протекают одновременно
Фотосинтез происходит в хлоропластах зеленых растений.
Фотосинтез начинается с улавливания и поглощения света пигментом хлорофиллом, содержащимся в хлоропластах клеток зеленых растений.
Этого
оказывается достаточно, чтобы сместить
спектр поглощения молекулы.

Молекула хлорофилла поглощает фотоны в фиолетовой и синей, а затем в красной части спектра, и не взаимодействует с фотонами в зеленой и желтой части спектра.
Поэтому хлорофилл и растения выглядят зелеными – они попросту никак не могут воспользоваться зелеными лучами и оставляют их гулять по белу свету (делая его тем самым зеленее).

Пигменты фотосинтеза располагаются на внутренней стороне мембраны тилакоидов.
Пигменты организованы в фотосистемы (антенные поля по улавливанию света) – содержащие по 250–400 молекул разных пигментов.
Фотосистема состоит из:
реакционного центра фотосистемы (молекула хлорофилла а),
антенных молекул
Все пигменты в фотосистеме способны передавать друг другу энергию возбужденного состояния. Энергия фотона, поглощенная той или иной молекулой пигмента, переносится на соседнюю молекулу, пока не достигнет реакционного центра. Когда резонансная система реакционного центра переходит в возбужденное состояние, она передает два возбужденных электрона молекуле-акцептору и тем самым окисляется и приобретает положительный заряд.
У растений:
фотосистема 1 (максимум поглощения света на длине волны 700 нм - Р700)
фотосистема 2 (максимум поглощения света на длине волны 680 нм - Р680
Различия в оптимумах поглощения обусловлены небольшими различиями в структуре пигментов.
Две системы работают сопряженно, как конвейер, состоящий из двух частей и называющийся нециклическим фотофосфорилированием .
Суммарное уравнение для нециклического фотофосфорилирования :
Ф - условное обозначение остатка фосфорной кислоты

Цикл начинается с фотосистемы 2.
1) антенные молекулы улавливают фотон и передают возбуждение молекуле активного центра Р680;
2) возбужденная молекула Р680 отдает два электрона кофактору Q при этом она окисляется и приобретает положительный заряд;
Кофактор (cofactor). Кофермент или любое другое вещество, необходимое для выполнения ферментом его функции
Коферменты (коэнзимы) [от лат. co (cum) - вместе и ферменты], органические соединения небелковой природы, участвующие в ферментативной реакции в качестве акцепторов отдельных атомов или атомных групп, отщепляемых ферментом от молекулы субстрата, т.е. для осуществления каталитического действия ферментов. Эти веществава, в отличие от белкового компонента фермента (апофермента), имеют сравнительно небольшую молекулярную массу и, как правило, термостабильны. Иногда под Коферментами подразумевают любые низкомолекулярные вещества, участие которых необходимо для проявления каталитического действия фермента, в т. ч. и ионы, напр. К + , Mg 2+ и Мn 2+ . Располагаются оферменты. в активном центре фермента и вместе с субстратом и функциональными группами активного центра образуют активированный комплекс.
Для проявления каталитической активности большинству ферментов необходимо наличие кофермента. Исключение составляют гидролитические ферменты (например, протеазы, липазы, рибонуклеаза), выполняющие свою функцию в отсутствие кофермента.
Молекула восстанавливается Р680 (под действием ферментов),. При этом вода диссоциирует на протоны и молекулярный кислород, т.е. вода является донором электронов, который обеспечивает восполнение электронов в Р 680.
ФОТОЛИЗ ВОДЫ - расщепление молекулы воды, в частности в процессе фотосинтеза. Вследствие фотолиза воды образуется кислород, выделяющийся зелеными растениями на свету.
История открытия удивительного и такого жизненного важного явления, как фотосинтез уходит корнями глубоко в прошлое. Более четырех веков назад в 1600 году бельгийский ученый Ян Ван - Гельмонт поставил простейший эксперимент. Он поместил веточку ивы в мешок, где находилось 80 кг земли. Ученый зафиксировал первоначальный вес ивы, и затем на протяжении пяти лет поливал растение исключительно дождевой водой. Каково же было удивление Яна Ван - Гельмонта, когда он повторно взвесил иву. Вес растения увеличился на 65 кг, причем масса земли уменьшился всего на 50 гр! Откуда растение взяло 64 кг 950 гр питательных веществ для ученого осталось загадкой!
Следующий значимый эксперимент на пути открытия фотосинтеза принадлежал английскому химику Джозефу Пристли. Ученый посадил под колпак мышь, и через пять часов грызун умер. Когда же Пристли поместил с мышью веточку мяты и также накрыл грызуна колпаком, мышь осталась живой. Этот эксперимент навел ученого на мысль о том, что существует процесс, противоположный дыханию. Ян Ингенхауз в 1779 году установил тот факт, что только зеленые части растений способны выделять кислород. Через три года швейцарский ученый Жан Сенебье доказал, что углекислый газ, под воздействием солнечных лучей, разлагается в зеленых органоидах растений. Спустя всего пять лет французский ученый Жак Буссенго, проводя лабораторные исследования, обнаружил тот факт, что поглощение растениями воды также происходит и при синтезе органических веществ. Эпохальное открытие в 1864 году совершил немецкий ботаник Юлиус Сакс. Ему удалось доказать, что объем потребляемого углекислого газа и выделяемого кислорода происходит в пропорции1:1.
Фотосинтез - один из самых значимых биологических процессов

Говоря научным языком, фотосинтез (от др.-греч. φῶς — свет и σύνθεσις — соединение, связывание) — это процесс, при котором из углекислого газа и воды на свету образуются органические вещества. Заглавная роль в этом процессе принадлежит фотосинтетическим сегментам.
Если говорить образно, то лист растения можно сравнить лабораторией, окна которой выходят на солнечную сторону. Именно в ней происходит образование органических веществ. Этот процесс является основой существования всего живого на Земле.
Многие резонно зададут вопрос: чем дышат люди, живущие в городе, где не то что дерева, и травинки днем с огнем не сыщешь. Ответ очень прост. Дело в том, что на долю наземных растений приходится всего 20% выделяемого растениями кислорода. Главенствующую роль в выработке кислорода в атмосферу играют морские водоросли. На их долю приходится 80% от вырабатываемого кислорода. Говоря языком цифр, и растения, и водоросли ежегодно выделяют в атмосферу 145 млрд. тонн (!) кислорода! Недаром мировой океан называют «легкими планеты».
Общая формула фотосинтеза выглядит следующим образом:
Вода + Углекислый газ + Свет → Углеводы + Кислород
Для чего нужен фотосинтез растениям?

Как мы уяснили, фотосинтез - это необходимое условие существования человека на Земле. Однако это не единственная причина, по которой фотосинтезирующие организмы производят активную выработку кислорода в атмосферу. Дело в том, что и водоросли, и растения ежегодно образуют более 100 млрд. органических веществ (!), которые составляют основу их жизнедеятельности. Вспоминая эксперимент Яна Ван-Гельмонта мы понимаем, что фотосинтез - это основа питания растений. Научно доказано, что 95% урожая определяют органические вещества, полученные растением в процессе фотосинтеза, и 5% - те минеральные удобрения, которые садовод вносит в почву.
Современные дачники основное внимание уделяют почвенному питанию растений, забывая о его воздушном питании. Неизвестно, какой урожай могли бы получить садоводы, если бы они внимательно относились к процессу фотосинтеза.
Однако ни растения, ни водоросли не могли бы так активно производить кислород и углеводы, не будь у них удивительного зеленого пигмента - хлорофилла.
Тайна зеленого пигмента

Главное отличие клеток растения от клеток иных живых организмов - это наличие хлорофилла. К слову сказать, именно он является виновником того, что листья растений окрашены именно в зеленый цвет. Это сложное органическое соединение обладает одним удивительным свойством: оно способно поглощать солнечный свет! Благодаря хлорофиллу становится возможны и процесс фотосинтеза.
Две стадии фотосинтеза
Говоря простым языком, фотосинтез представляет собой процесс, при котором поглощенные растением вода и углекислый газ на свету при помощи хлорофилла образуют сахар и кислород. Таким образом, неорганические вещества удивительным образом превращаются в органические. Полученный в результате преобразования сахар является источником энергии растений.

Фотосинтез имеет две стадии: световую и темновую.
Световая фаза фотосинтеза
Осуществляется на мембранах тилакойдов.
Тилакойд - это структуры, ограниченные мембраной. Они располагаются в строме хлоропласта.
Порядок событий световой стадии фотосинтеза:
- На молекулу хлорофилла попадает свет, который затем поглощается зеленым пигментом и приводит его в возбужденное состояние. Входящий в состав молекулы электрон переходит на более высокий уровень, участвует в процессе синтеза.
- Происходит расщепление воды, в ходе которого протоны под воздействием электронов превращаются в атомы водорода. Впоследствии они расходуются на синтез углеводов.
- На завершающем этапе световой стадии происходит синтез АТФ (Аденозинтрифосфат). Это органическое вещество, которое играет роль универсального аккумулятора энергии в биологических системах.
Темновая фаза фотосинтеза
Местом протекания темновой фазы являются строму хлоропластов. Именно в ходе темновой фазы происходит выделение кислорода и синтез глюкозы. Многие подумают, что такое название эта фаза получила потому что процесс, происходящие в рамках этого этапа осуществляются исключительно в ночное время. На самом деле, это не совсем верно. Синтез глюкозы происходит круглосуточно. Дело в том, что именно на данном этапе световая энергия больше не расходуется, а значит, она попросту не нужна.
Значение фотосинтеза для растений

Мы уже определили тот факт, что фотоинтез нужен растениям ничем не меньше, чем нам. О масштабах фотосинтеза очень просто говорить языком цифр. Ученые рассчитали, что только растения суши запасают столько солнечной энергии, сколько могли бы израсходовать 100 мегаполисов в течение 100 лет!
Дыхание растений - это процесс, противоположный фотосинтезу. Смысл дыхания растений заключается в освобождении энергии в процессе фотосинтеза и направление ее на нужды растений. Говоря простым языком, урожай - это разница между фотосинтезом и дыханием. Чем больше фотосинтез и ниже дыхание, тем больше урожай, и наоборот!
Фотосинтез - это удивительный процесс, который делает возможной жизнь на Земле!
Любой зеленый листик – это миниатюрная фабрика питательных веществ и кислорода, который необходим животным и человеку для нормальной жизнедеятельности. Процесс выработки данных веществ из воды и углекислоты из атмосферы называют фотосинтезом. Фотосинтез – это сложнейший химический процесс, который происходит с участием света. Конечно же, всем интересно как происходит фотосинтез. Сам процесс состоит из двух этапов: первый - это поглощение квантов света, а второй - использование их энергии в разных химических реакциях.
Как происходит процесс фотосинтеза
Растение поглощает свет при помощи зеленого вещества, которое называется хлорофилл. Хлорофилл содержится в хлоропластах, которые находятся в стеблях или плодах. Особенно большое их количество в листьях, потому что из-за своей очень плоской структуры листок может притянуть много света, соответственно, получить намного больше энергии для процесса фотосинтеза.
После поглощения хлорофилл находится в возбужденном состоянии и передает энергию другим молекулам организма растения, особенно, тем, которые непосредственно участвуют в фотосинтезе. Второй этап процесса фотосинтеза проходит уже без обязательного участия света и состоит в получении химической связи с участием углекислого газа, получаемого из воздуха и воды. На данной стадии синтезируются разные очень полезные для жизнедеятельности вещества, такие как крахмал и глюкоза.
Эти органические вещества используют сами растения для питания разных его частей, а также для поддержания нормальной жизнедеятельности. Кроме того, эти вещества также получают и животные, питаясь растениями. Люди тоже получают эти вещества, употребляя в пищу продукты животного и растительного происхождения.
Условия для фотосинтеза
Фотосинтез может происходить как под действием искусственного света, так и солнечного. Как правило, на природе растения интенсивно «работают» в весенне-летний период, когда необходимого солнечного света много. Осенью света меньше, день укорачивается, листья сначала желтеют, а потом опадают. Но стоит появиться весеннему теплому солнцу, как зеленая листва вновь появляется и зеленые «фабрики» снова возобновят свою работу, чтобы давать кислород, такой необходимый для жизни, а также множество других питательных веществ.
Где происходит фотосинтез
В основном фотосинтез, как процесс, происходит, как это уже было сказано, в листьях растений, потому как они способны принять на себя больше солнечного света, который очень необходим для процесса фотосинтеза.
Как итог можно сказать то, что процесс фотосинтеза является неотъемлемой частью жизнедеятельности растений.
5. Фазы и процессы фотосинтеза (продолжение)Рассмотренные нами в предыдущих частях два этапа фотосинтеза – физический и фотохимический – объединяют в так называемую световую фазу фотосинтеза. Теперь же речь пойдет о второй фазе фотосинтеза, часто называемой темновой фазой.
Темновая фаза – не совсем удачное название. Если мы говорим, что световая фаза названа так, чтобы подчеркнуть зависимость всех протекающих в ней реакций от света, то название «темновая фаза» означает, что все реакции, в ней протекающие, от света не зависят и проходят в темноте.
Но это не совсем точно. Очень многие реакции темновой фазы фотосинтеза зависят от света, так как ферменты, катализирующие эти реакции, индуцируются светом. Поэтому эту фазу фотосинтеза лучше называть путем превращения углерода, или циклом фиксации углерода, – по основному процессу, который в ней происходит.
(Здесь нужно обязательно пояснить, что происходит фиксация не чистого углерода, а углерода в составе углекислого газа СО 2 .)
Отметим, что разделение процесса фотосинтеза на две фазы происходит не только по отношению к свету, но и по месту протекания реакций. Реакции световой фазы протекают в тилакоидах гран и стромы, а реакции фиксации углерода протекают в матриксе (строме) хлоропластов.
Стоит обратить внимание учащихся на то, что в литературе встречается и другое название тилакоидов – ламеллы гран. Взаимосвязь терминов можно объяснить, зачитав отрывок из «Физиологии растений» Н.И. Якушкина: «Внутреннее пространство хлоропластов заполнено бесцветным содержимым – стромой – и пронизано мембранами (ламеллами). Ламеллы, соединенные друг с другом, образуют как бы пузырьки – тилакоиды. В хлоропластах тилакоиды двух типов. Короткие тилакоиды собраны в пачки и расположены друг над другом, напоминая стопку монет. Эти стопки называются гранами, а составляющие их ламеллы – ламеллами гран. Между гранами параллельно друг другу располагаются длинные тилакоиды. Составляющие их ламеллы получили название ламеллы стром».
Рассматривая Z-схему, мы установили, что конечные продукты циклического и нециклического фосфорилирования – АТФ и НАДФ . Н – используются в темновых реакциях фотосинтеза. Как же они используются?
Если в световой фазе АТФ и НАДФ . Н являются конечными продуктами, то в процессе фиксации углерода они используются на самом первом этапе всего цикла фиксации углерода. Весь цикл фиксации углерода можно представить в виде следующих стадий.
Первая стадия – непосредственная фиксация углекислого газа – карбоксилирование.
Вторая стадия – образование 3-фосфоглицеринового альдегида (ФГА).
Третья стадия – образование продуктов фотосинтеза.
Четвертая стадия – восстановление первоначальных реагентов.
Перечисленные стадии выделены условно – вместе они составляют цикл фиксации углерода, или цикл Кальвина.
В отличие от световых реакций, которые протекали в строгой последовательности, реакции фиксации углерода могут протекать параллельно, за исключением первых двух – фиксации углекислого газа и образования ФГА. Рассмотрим каждую стадию цикла.
КарбоксилированиеЭта стадия – ключевая, потому что в ней участвует СО 2 . Молекула углекислого газа соединяется с молекулой пятиуглеродного сахара рибулезодифосфата (РДФ) с образованием нестойкого шестиуглеродного соединения, которое затем распадается на две молекулы 3-фосфоглицериновой кислоты (1).
Реакция карбоксилирования очень интересна тем, что в зависимости от условий она может протекать с образованием различных конечных продуктов. Так, например, при наличии СО 2 продуктом реакции будет только ФГК, а в присутствии О 2 РДФ не присоединяет углекислый газ и распадается на ФГК и фосфогликолевую кислоту, которая используется в процессах фотодыхания. Фотодыхание – это процесс, протекающий лишь на свету и сопровождающийся поглощением О 2 и выделением СО 2 . Такое изменение хода реакции объясняется тем, что участвующий в ней фермент обладает двойной каталитической активностью – по отношению к углегислому газу и кислороду.
Этот фермент называется рибулозо-1,5-бифосфаткарбоксилаза-оксигеназа (РуБФ-карбоксилаза). Этот фермент составляет около 50% всех растворимых белков в листьях и потому может считаться самым распространенным белком в природе. Фермент состоит из двух субъединиц – большой и малой. Интересно, что белки больших субъединиц кодируются ДНК хлоропластов, а белки малых субъединиц – ядерной ДНК. Большие субъединцы обладают каталитической активностью и в отсутствие малых, которые, по-видимому, играют регуляторную роль. Этот факт может служить подтверждением того, что хлоропласты произошли от прокариотических предков.
Таким образом, на первых этапах фиксации углерода имеет место конкуренция между двумя процессами – фиксацией углерода и фотодыханием. Для сдвига баланса в сторону фиксации углерода необходимы ионы Мg 2 + (2).
Образование фосфоглицеринового альдегидаОбразующаяся на первой стадии ФГК превращается в ФГА в два этапа (3 и 4). Сначала используется АТФ, синтезированная в световой фазе фотосинтеза. Затем используется НАДФ . Н, который тоже является продуктом световой фазы фотосинтеза.
Молекула ФГА является ключевым веществом для третьей стадии.
Образование продуктов фотосинтезаОбычно продуктом фотосинтеза называют сахар. На самом деле продуктами фотосинтеза можно считать и другие вещества, о чем мы упоминали при рассмотрениии Z-схемы.
Молекула ФГА используется растением в цикле Кальвина в нескольких направлениях.
Во-первых, ФГА является основой для синтеза сахара.
Во-вторых, ФГА может быть
использована для синтеза аминокислот.
Среди продуктов фотосинтеза обнаружены такие
аминокислоты, как аланин, серин, глютаминовая
кислота, глицин. Синтез аминокислот происходит
интенсивно при недостатке НАДФ .
Н, в
результате чего из ФГК образуется не ФГА, а
пировиноградная кислота, которая является
исходным соединением для синтеза аминокислот и
одним из ключевых веществ цикла Кребса.
В-третьих, ФГА дает начало циклу превращений некоторых промежуточных продуктов в РДФ, который служит акцептором углекислого газа.
Наряду с углеводами и аминокислотами из промежуточных продуктов цикла Кальвина могут образовываться липиды и другие продукты.
Во всех уравнениях фотосинтеза в правой части пишется формула шестиуглеродного сахара. Как правило, его называют глюкозой. Но в действительности первым свободным сахаром является дисахарид сахароза, из которой образуются два моносахарида – глюкоза и фруктоза.
Восстановление первоначальных реагентовДля того чтобы растение могло акцептировать новую молекулу углекислого газа, необходимо иметь РДФ, основной акцептор углекислого газа. РДФ образуется из ФГА в результате цепи реакций, в процессе которых образуются пяти- и семиуглеродные сахара. Надо отметить, что основная масса ФГА идет именно на восстановление нужного количества РДФ: из 12 образовавшихся молекул ФГА только две идут на образование продуктов фотосинтеза, т.е. сахарозы.
Подводя итог рассмотрению фаз фотосинтеза, можно составить обобщенную схему фотосинтеза (рис. 1).
Учитывая реакции световой и темновой фаз фотосинтеза, можно привести следующее суммарное уравнение фотосинтеза.
Световые реакции:
Темновые реакции:
6. Виды фотосинтеза
В настоящее время известны три разных механизма темновых реакций фотосинтеза у высших растений. Но, по-видимому, правильнее говорить об одном основном процессе и двух вариантах.
Основной механизм – это фиксация углерода в цикле Кальвина. В последнее время этот цикл стали называть С 3 -путем, или С 3 -типом, фотосинтеза, а растения, осуществляющие только реакции этого цикла, называют С 3 -растениями. Такие растения обычно растут в областях умеренного климата; оптимальная дневная температура для фиксации углекислого газа у этих растений составляет от +15 до +25 °С.
Первый вариант – это С 4 -путь (или С 4 -тип фотосинтеза), называемый также циклом Хетча–Слэка. Растения, осуществляющие данный тип фотосинтеза, распространены в тропических и субтропических областях.
Второй вариант – процесс, известный под названием метаболизма органических кислот по типу толстянковых (МОКТ- или САМ-фотосинтез). Растения с таким типом фотосинтеза часто встречаются в засушливых пустынных областях.
С 3 -растения превращают СО 2 в углеводы только в реакциях цикла Кальвина. С 4 -растения и МОКТ-растения также осуществляют цикл Кальвина, но в них поглощение СО 2 и превращение его в углеводы включает в себя и другие реакции. С 4 -растения и МОКТ отличаются друг от друга природой этих дополнительных реакций, временем суток, когда они происходят, и тем, в каких клетках находятся вещества, участвующие в этих реакциях.
У С 3 -растений фотосинтез происходит только в клетках мезофилла листа, а у С 4 -растений – в клетках мезофилла и в клетках обкладки сосудистых пучков.
С4-тип фотосинтезаВ самых общих чертах путь углерода в реакциях С 4 -типа фотосинтеза показан на рис. 2.
Рис. 2. Схематическое изображение пути углерода при С 4 -фотосинтезе. С 3 -соединения содержат три атома углерода в молекуле, С 4 -соединения – четыре
Цикл Кальвина у данного типа растений осуществляется в клетках обкладки сосудистого пучка и протекает так же, как у С 3 -растений.
Фиксация углекислого газа у С 3 - и С 4 -растений значительно различается. Если у С 3 -растений молекула углекислого газа присоединялась к пятиуглеродной молекуле РДФ, то у С 4 -растений акцептором углекислого газа является трехуглеродная молекула, чаще всего – это фосфоенолпировиноградная кислота (ФЕП). Соединяясь с углекислым газом ФЕП превращается в щавелевоуксусную кислоту (ЩУК), которая и поступает в хлоропласт клеток мезофилла. В хлоропластах ЩУК при наличии НАДФ. Н превращается в яблочную кислоту (ЯК), которая поступает в клетки обкладки сосудистых пучков. В клетках обкладки сосудистых пучков ЯК отдает молекулу углекислого газа в цикл Кальвина, превращаясь в пировиноградную кислоту (ПВК). ПВК, в свою очередь, возвращается в хлоропласты мезофилла, превращается в ФЕП, и начинается новый цикл (рис. 3).
Рис. 3. Фотосинтез С4-типа (на примере кукурузы)
Увеличение числа реакций для фиксации углекислого газа у С 4 -растений на первый взгляд может показаться излишним и бессмысленным. Но это только на первый взгляд. Растениям с С 4 -типом фотосинтеза приходится концентрировать углекислый газ в клетках обкладки, т.к. по сравнению с С 3 -растениями в их клетках углекислого газа содержится значительно меньше. Это связано с тем, что С 4 -растения обитают в более жарком и сухом климате, чем С 3 -растения, поэтому для уменьшения потерь воды им приходится уменьшать транспирацию. За счет этого создаются трудности в поглощении углекислого газа, что и приводит к необходимости его концентрации. В настоящее время считается, что С 4 -тип фотосинтеза является эволюционным приспособлением к более жарким и сухим климатическим условиям.
Метаболизм органических кислот по типу толстянковых (МОКТ)Растения с данным типом фотосинтеза являются в основном суккулентами.
Для МОКТ-растений характерны следующие особенности.
1. Их устьица обычно открыты ночью (т.е. в темноте) и закрыты в течение дня.
2. Фиксация углекислого газа происходит в темное время суток. При этом образуется значительное количество яблочной кислоты.
3. Яблочная кислота запасается в больших вакуолях, которые характерны для клеток МОКТ-растений.
4. В светлое время суток яблочная кислота отдает углекислый газ в цикл Кальвина, где она превращается в сахарозу или запасной углевод глюкан.
5. В темновой период суток часть запасенного глюкана распадается с образованием молекул-акцепторов для темновой фиксации углекислого газа (рис. 4).
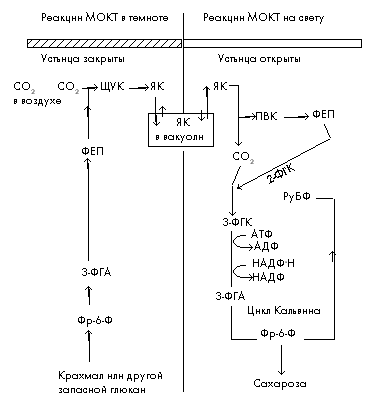
Таким образом у МОКТ-растений существует суточный ритм: ночью содержание запасного глюкана падает и содержание яблочной кислоты повышается, а днем происходят противоположные изменения.
В заключение надо добавить, что фотосинтез по МОКТ-типу считается самым поздним приспособлением растений в процессе эволюции.
Осуществляется процесс фотосинтеза в листьях растений. Фотосинтез свойствен лишь зеленым растениям. Эту важнейшую сторону деятельности листа полнее всего характеризует К. А. Тимирязев:
Можно сказать, что в жизни листа выражается самая сущность растительной жизни. Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались - в растении ли, в животном или в человеке, - прошли через лист, произошли из веществ, выработанных листом.
Строение листьев растений
Листья растений по анатомическому строению отличаются большим разнообразием, которое зависит и от вида растения, и от условий их роста. Лист сверху и снизу покрыт эпидермисом - покровной тканью с многочисленными отверстиями, называемыми устьицами. Под верхним эпидермисом расположена палисадная, или столбчатая паренхима, называемая ассимиляционной. Под ней находится более рыхлая ткань - губчатая паренхима, за которой идет нижний эпидермис. Весь лист пронизан сетью жилок, состоящих из проводящих пучков, по которым проходят вода, минеральные и органические вещества. Поперечный разрез листа. В столбчатой и губчатой ткани листа расположены зеленые пластиды - хлоропласты, содержащие пигменты. Наличием хлоропластов и содержащихся в них зеленых пигментов (хлорофиллов) объясняется окраска растений. Огромная листовая поверхность, достигающая 30 000 - 50 000 кв. м на 1 га у разных растений, хорошо приспособлена для успешного поглощения СО 2 из воздуха в процессе фотосинтеза. Углекислый газ проникает в лист растения через устьица, расположенные в эпидермисе, поступает в межклетники и, проникая через оболочку клеток, попадает в цитоплазму, а затем в хлоропласты, где и осуществляется процесс ассимиляции. Образующийся в этом процессе кислород диффундирует с поверхности хлоропластов в свободном состоянии. Таким образом, через устьица осуществляется газообмен листьев с внешней средой - поступление углекислого газа и выделение кислорода в процессе фотосинтеза, выделение углекислого газа и поглощение кислорода в процессе дыхания. Кроме того, устьица служат для выделения паров воды. Несмотря на то, что общая площадь устьичных отверстий составляет лишь 1-2% всей листовой поверхности, тем не менее при открытых устьицах углекислый газ проникает в листья со скоростью, превышающей в 50 раз поглощение его щелочью. Количество устьиц очень велико - от нескольких десятков до 1500 на 1 кв. мм.Хлоропласты
Хлоропласты - зеленые пластиды, в которых происходит процесс фотосинтеза. Они расположены в цитоплазме. У высших растений хлоропласты имеют дискообразную или линзовидную форму, у низших они более разнообразны. Хлоропласты в клетках зеленых растений.
Размер хлоропластов у высших растений довольно постоянен, составляя в среднем 1 -10 мк. Обычно в клетке содержится большое количество хлоропластов, в среднем 20-50, а иногда и больше. Расположены они главным образом в листьях, много их в незрелых плодах. В растении общее количество хлоропластов огромно; во взрослом дереве дуба, например, площадь их равняется 2 га.
Хлоропласт имеет мембранную структуру. От цитоплазмы он отделен двухмембранной оболочкой. В хлоропласте находятся ламеллы, белково-липоидные пластинки, собранные в пучки и называемые гранами.
Хлорофилл расположен в ламеллах в виде мономолекулярного слоя. Между ламеллами находится водянистая белковая жидкость - строма; в ней встречаются крахмальные зерна и капли масла.
Строение хлоропласта хорошо приспособлено к фотосинтезу, так как разделение хлорофиллоносного аппарата на мелкие пластинки значительно увеличивает активную поверхность хлоропласта, что облегчает доступ энергии и перенос ее к химическим системам, участвующим в фотосинтезе.
Данные А. А. Табенцкого показывают, что хлоропласты все время изменяются в онтогенезе растения. В молодых листьях наблюдается мелкогранулярная структура хлоропластов, в листьях, закончивших рост,- крупногранулярная.
В старых листьях уже наблюдается распад хлоропластов. В сухом веществе хлоропластов содержится 20-45% белков, 20-40% липоидов, 10-12% углеводов и других запасных веществ, 10% минеральных элементов, 5-10% зеленых пигментов (хлорофилл а
и хлорофилл б
), 1-2% каротиноидов, а также небольшое количество РНК и ДНК. Содержание воды достигает 75%.
В хлоропластах имеется большой набор гидролитических и окислительно-восстановительных ферментов. Исследованиями Н. М. Сисакяна показано, что в хлоропластах происходит и синтез многих ферментов. Благодаря этому они принимают участие во всем сложном комплексе процессов жизнедеятельности растения.
Хлоропласты в клетках зеленых растений.
Размер хлоропластов у высших растений довольно постоянен, составляя в среднем 1 -10 мк. Обычно в клетке содержится большое количество хлоропластов, в среднем 20-50, а иногда и больше. Расположены они главным образом в листьях, много их в незрелых плодах. В растении общее количество хлоропластов огромно; во взрослом дереве дуба, например, площадь их равняется 2 га.
Хлоропласт имеет мембранную структуру. От цитоплазмы он отделен двухмембранной оболочкой. В хлоропласте находятся ламеллы, белково-липоидные пластинки, собранные в пучки и называемые гранами.
Хлорофилл расположен в ламеллах в виде мономолекулярного слоя. Между ламеллами находится водянистая белковая жидкость - строма; в ней встречаются крахмальные зерна и капли масла.
Строение хлоропласта хорошо приспособлено к фотосинтезу, так как разделение хлорофиллоносного аппарата на мелкие пластинки значительно увеличивает активную поверхность хлоропласта, что облегчает доступ энергии и перенос ее к химическим системам, участвующим в фотосинтезе.
Данные А. А. Табенцкого показывают, что хлоропласты все время изменяются в онтогенезе растения. В молодых листьях наблюдается мелкогранулярная структура хлоропластов, в листьях, закончивших рост,- крупногранулярная.
В старых листьях уже наблюдается распад хлоропластов. В сухом веществе хлоропластов содержится 20-45% белков, 20-40% липоидов, 10-12% углеводов и других запасных веществ, 10% минеральных элементов, 5-10% зеленых пигментов (хлорофилл а
и хлорофилл б
), 1-2% каротиноидов, а также небольшое количество РНК и ДНК. Содержание воды достигает 75%.
В хлоропластах имеется большой набор гидролитических и окислительно-восстановительных ферментов. Исследованиями Н. М. Сисакяна показано, что в хлоропластах происходит и синтез многих ферментов. Благодаря этому они принимают участие во всем сложном комплексе процессов жизнедеятельности растения.
Пигменты, их свойства и условия образования
Пигменты можно извлечь из листьев растений спиртом или ацетоном. В вытяжке находятся следующие пигменты: зеленые - хлорофилл а и хлорофилл б ; желтые - каротин и ксантофилл (каротиноиды).Хлорофилл
Хлорофилл представляет собойодно из интереснейших веществ на земной поверхности(Ч. Дарвин), так как благодаря ему возможен синтез органических веществ из неорганических СО 2 и Н 2 О. Хлорофилл не растворяется в воде, под влиянием солей, кислот и щелочей легко изменяется, поэтому было очень трудно установить его химический состав. Для извлечения хлорофилла обычно применяют этиловый спирт или ацетон. Хлорофилл имеет следующие суммарные формулы: хлорофилл а - С 55 Н 72 О 5 N 4 Mg, хлорофилл б - С 55 Н 70 О 6 N 4 Mg. У хлорофилла а больше на 2 атома водорода и меньше на 1 атом кислорода, чем у хлорофилла б . Формулы хлорофилла можно представить и так:
 Формулы хлорофилла а
и б
.
Центральное место в молекуле хлорофилла занимает Мg; его можно вытеснить, подействовав на спиртовую вытяжку хлорофилла соляной кислотой. Зеленый пигмент превращается в бурый, называемый феофитином, в котором Мg замещается двумя атомами Н из соляной кислоты.
Восстановить зеленый цвет вытяжки очень легко внесением в молекулу феофитина магния или другого металла. Следовательно, зеленый цвет хлорофилла связан с наличием в его составе металла.
При воздействии на спиртовую вытяжку хлорофилла щелочью происходит отщепление спиртовых групп (фитола и метилового спирта); в этом случае зеленая окраска хлорофилла сохраняется, что указывает на сохранение ядра молекулы хлорофилла при этой реакции.
Химический состав хлорофилла у всех растений одинаков. Содержание хлорофилла а всегда больше (примерно в 3 раза), чем хлорофилла б. Общее количество хлорофилла невелико и составляет около 1 % от сухого вещества листа.
По своей химической природе хлорофилл близок к красящему веществу крови - гемоглобину, центральное место в молекуле которого занимает не магний, а железо. В соответствии с этим различаются и их физиологические функции: хлорофилл принимает участие в важнейшем восстановительном процессе в растении - фотосинтезе, а гемоглобин - в процессе дыхания животных организмов, перенося кислород.
Формулы хлорофилла а
и б
.
Центральное место в молекуле хлорофилла занимает Мg; его можно вытеснить, подействовав на спиртовую вытяжку хлорофилла соляной кислотой. Зеленый пигмент превращается в бурый, называемый феофитином, в котором Мg замещается двумя атомами Н из соляной кислоты.
Восстановить зеленый цвет вытяжки очень легко внесением в молекулу феофитина магния или другого металла. Следовательно, зеленый цвет хлорофилла связан с наличием в его составе металла.
При воздействии на спиртовую вытяжку хлорофилла щелочью происходит отщепление спиртовых групп (фитола и метилового спирта); в этом случае зеленая окраска хлорофилла сохраняется, что указывает на сохранение ядра молекулы хлорофилла при этой реакции.
Химический состав хлорофилла у всех растений одинаков. Содержание хлорофилла а всегда больше (примерно в 3 раза), чем хлорофилла б. Общее количество хлорофилла невелико и составляет около 1 % от сухого вещества листа.
По своей химической природе хлорофилл близок к красящему веществу крови - гемоглобину, центральное место в молекуле которого занимает не магний, а железо. В соответствии с этим различаются и их физиологические функции: хлорофилл принимает участие в важнейшем восстановительном процессе в растении - фотосинтезе, а гемоглобин - в процессе дыхания животных организмов, перенося кислород.
Оптические свойства пигментов
Хлорофилл поглощает солнечную энергию и направляет ее на химические реакции, которые не могут протекать без энергии, получаемой извне. Раствор хлорофилла в проходящем свете имеет зеленый цвет, но при увеличении толщины слоя или концентрации хлорофилла он приобретает красный цвет. Хлорофилл поглощает свет не сплошь, а избирательно. При пропускании белого света через призму получается спектр, состоящий из семи видимых цветов, которые постепенно переходят друг в друга. При пропускании белого света через призму и раствор хлорофилла на полученном спектре наиболее интенсивное поглощение будет в красных и сине-фиолетовых лучах. Зеленые лучи поглощаются мало, поэтому в тонком слое хлорофилл имеет в проходящем свете зеленый цвет. Однако с увеличением концентрации хлорофилла полосы поглощения расширяются (значительная часть зеленых лучей также поглощается) и без поглощения проходит только часть крайних красных. Спектры поглощения хлорофилла а и б очень близки. В отраженном свете хлорофилл кажется вишнево-красным, так как он излучает поглощенный свет с изменением длины его волны. Это свойство хлорофилла называется флюоресценцией.Каротин и ксантофилл
Каротин и ксантофилл имеют полосы поглощения только в синих и фиолетовых лучах. Их спектры близки друг другу. Спектры поглощения хлорофиллом а
и б
.
Поглощенная этими пигментами энергия передается хлорофиллу а
, который является непосредственным участником фотосинтеза. Каротин считают провитамином А, так как при его расщеплении образуются 2 молекулы витамина А. Формула каротина - С 40 Н 56 , ксантофилла - С 40 Н 54 (ОН) 2 .
Спектры поглощения хлорофиллом а
и б
.
Поглощенная этими пигментами энергия передается хлорофиллу а
, который является непосредственным участником фотосинтеза. Каротин считают провитамином А, так как при его расщеплении образуются 2 молекулы витамина А. Формула каротина - С 40 Н 56 , ксантофилла - С 40 Н 54 (ОН) 2 .
Условия образования хлорофилла
Образование хлорофилла осуществляется в 2 фазы: первая фаза - темновая, во время которой образуется предшественник хлорофилла - протохлорофилл, а вторая - световая, при которой из протохлорофилла на свету образуется хлорофилл. Образование хлорофилла зависит как от вида растения, так и от ряда внешних условий. Некоторые растения, например проростки хвойных, могут позеленеть и без участия света, в темноте, но у большинства растений хлорофилл образуется из протохлорофилла только на свету. В отсутствие света получаются этиолированные растения, имеющие тонкий, слабый, сильно вытянутый стебель и очень мелкие бледно-желтые листья. Если выставить этиолированные растения на свет, то листья быстро позеленеют. Это объясняется тем, что в листьях уже имеется протохлорофилл, который под воздействием света легко превращается в хлорофилл. Большое влияние на образование хлорофилла оказывает температура; при холодной весне у некоторых кустарников листья не зеленеют до установления теплой погоды: при понижении температуры подавляется образование протохлорофилла. Минимальной температурой, при которой начинается образование хлорофилла, является 2°, максимальной, при которой образование хлорофилла не происходит, 40°. Кроме определенной температуры, для образования хлорофилла необходимы элементы минерального питания, особенно железо. При его отсутствии у растений наблюдается заболевание, называемое хлорозом. По-видимому, железо является катализатором при синтезе протохлорофилла, так как в состав молекулы хлорофилла оно не входит. Для образования хлорофилла также необходимы азот и магний, входящие в состав его молекулы. Важным условием является и наличие в клетках листа пластид, способных к позеленению. При их отсутствии листья растений остаются белыми, растение не способно к фотосинтезу и может жить только до тех пор, пока не израсходует запасы семени. Это явление называется альбинизмом. Оно связано с изменением наследственной природы данного растения.Количественные отношения между хлорофиллом и усваиваемой углекислотой
При большем содержании хлорофилла в растении процесс фотосинтеза начинается при меньшей интенсивности света и даже при более низкой температуре. С увеличением содержания хлорофилла в листьях фотосинтез возрастает, но до известного предела. Следовательно, нет прямой зависимости между содержанием хлорофилла и интенсивностью поглощения СО 2 . Количество ассимилированного листом СО 2 в час в пересчете на единицу содержащегося в листе хлорофилла тем выше, чем меньше хлорофилла. Р. Вильштеттером и А. Штолем была предложена единица, характеризующая соотношение между количеством хлорофилла и поглощенным углекислым газом. Количество разложенной в единицу времени углекислоты, приходящееся на единицу веса хлорофилла, они назвали ассимиляционным числом . Ассимиляционное число непостоянно: оно больше при малом содержании хлорофилла и меньше при высоком содержании его в листьях. Следовательно, молекула хлорофилла используется более продуктивно при низком его содержании в листе и продуктивность хлорофилла уменьшается с увеличением его количества. Данные введены в таблицу.Таблица Ассимиляционное число в зависимости от содержания хлорофилла (по Р. Вильштеттеру и А. Штолю)
| Растения |
в 10г. листьев (мг) |
Ассимиляционное число |
|
зеленая раса желтая раса |
16,2 1,2 | 6,9 82,0 |
| Сирень | 16,2 | 5,8 |
| Этиолированные проростки фасоли после освещения в течение: 6 часов 4 дней |


















